|
|
|||||||||||||||||||||
|
Como vimos en el
capítulo precedente desde sus inicios el desarrollo
del aparato genital se encuentra en íntima
relación con el desarrollo del aparato urinario.
Así, mientras el mesonefros es aún el
órgano excretor del embrión, la gónada
se manifiesta hacia finales de la quinta semana del
desarrollo como un engrosamiento del mesotelio y del
mesénquima subyacente, el pliegue gonadal, eminencia
que se ubica medial al mesonefros. |
|||||||||||||||||||||
|
|||||||||||||||||||||
|
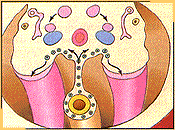
Hacia la cuarta
semana de desarrollo las células germinales
primordiales migran, mediante movimientos ameboideos, por el
mesenterio dorsal hacia la pared corporal posterior del
embrión en dirección al pliegue gonadal,
sufriendo sucesivas mitosis durante la migración lo
que permite incrementar su número. De modo que hacia
el final de la quinta semana alrededor del 2.500
células germinales primordiales comienzan a poblar
los pliegues genitales; pliegues que se encuentran al nivel
del décimo segmento vertebral torácico.
Algunas
células germinales primordiales pueden extraviar su
ruta de migración e implantarse en sitios
inapropiados. Ocasionalmente estas células
ectópicas pueden proliferar y dar
origen a un tipo particular de tumor llamado
teratoma.
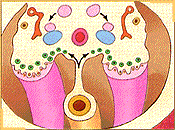
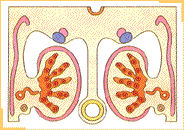
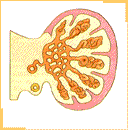
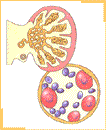
Durante la sexta semana de desarrollo la llegada de células germinales a la región de los pliegues gonadales estimula a las células celómicas y del mesénquima subyacente las que proliferan y forman bandas compactas de tejido, los cordones sexuales primitivos, los cuales rodean a las células germinales. En los pliegues gonadales, simétricamente dispuestos en el lado del mesenterio dorsal, es posible reconocer una región cortical y una región medular. El destino final de estas regiones va a depender del sexo genético del embrión. Si bien la presencia de las células germinales primordiales es necesaria para el desarrollo completo de la diferenciación gonadal, en los embriones de sexo masculino en ausencia de ellas se produce el desarrollo inicial del testículo. Hacia finales de la quinta e inicios de la sexta semana un nuevo par de conductos, los conductos paramesonéfricos (de Müller), comienzan a formarse lateralmente a los conductos mesonéfricos en embriones de ambos sexos.
Como podemos ver
las estructuras genitales internas presenten un estado
inicial indiferenciado, extendido entre la cuarta y la sexta
semana de desarrollo, que posteriormente va a definirse para
expresar las características de uno u otro
sexo.
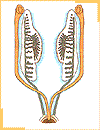
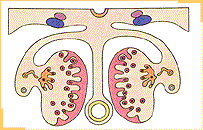
Desarrollo del testículoLa diferenciación del testículo precede a la diferenciación del ovario. Así, durante la sexta semana del desarrollo, bajo la influencia de la proteína SRY, los cordones sexuales primitivos se condensan hacia la región medular de la gónada formando los cordones sexuales medulares.En la pared de éstos se diferencian las células de Sertoli, mientras que se produce la involución de los cordones sexuales corticales.
La
porción más periférica de los cordones
sexuales medulares forman los túbulos
seminíferos; en cambio la zona profunda de ellos
adopta una disposición reticular formando la rete
testis (red de Haller).
De modo que hacia la décima semana los testículos sobresalen en la pared abdominal posterior del embrión, bajo el mesotelio (peritoneo primitivo), separados del mesonefros y unida a la pared corporal por el mesorquio.
Esta situación cambia después de la pubertad, momento en que comienza la espermatogénesis. Desarrollo del ovarioEn ausencia de factores virilizantes como la proteína SRY o la hormona antimülleriana el pliegue gonadal comienza a diferenciarse en ovario. En contraste con lo que ocurre con el testículo, la presencia de las células germinales primordiales es esencial para la diferenciación del ovario; si estas células no aparecen o son anormales, como ocurre en el síndrome de Turner (X0), la gónada se retrae y se forman ovarios vestigiales.
Los elementos celulares de estos cordones corticales rodean a las células germinales primordiales.
Muchas de estas ovogonias degeneran y al séptimo mes de vida intrauterina todas las ovogonias ingresan en su primera división meiótica, tomando el nombre ahora de ovocitos primarios; células germinales que se encuentran en el interior de los folículos primordiales del ovario fetal. El proceso de la primera división meiótica se detiene en la profase 1 al momento del nacimiento; proceso que será reasumido a partir de la pubertad. El ovario en desarrollo hace eminencia en la cavidad abdominal del feto y se mantiene unida a la pared corporal a través del mesovario. En la superficie del ovario fetal existe una delgada túnica fibrosa, la túnica albugínea, tejido que después se integra a la corteza del ovario. Los cordones sexuales medulares en el embrión de sexo masculino participan el la formación de la rete testis; en el feto femenino forman una red laxa, la rete ovarii, que posteriormente se desintegra. Del mismo modo, en los embriones femeninos el conducto mesonéfrico involuciona persistiendo algunos vestigios epiteliales que forman remanentes como el epooforo y el paraoforo, en relación con el mesoovario, y formaciones quísticas (quistes de Gartner) en relación con la vagina. |
|||||||||||||||||||||
|
[Introducción][Diferenciación de la gónada][Conductos genitales][Descenso gonadal][Genitales externos]
|