
|
|
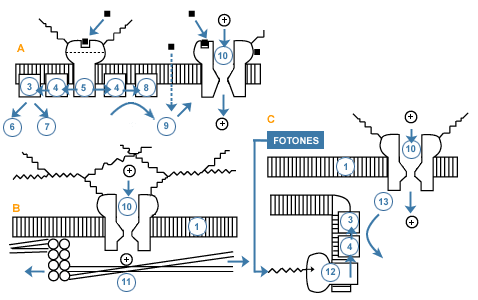
En el proceso de transducción la energía del estímulo es transformada en una señal bioeléctrica. A pesar de los diversos tipos de receptores que existen en un organismo es posible sin embargo, resumir en tres mecanismos básicos de transducción la aparente diversidad que para este proceso podría encontrarse en los receptores sensoriales. En los tres modelos se llega, con participación en algunos de ellos de segundos mensajeros, a la modificación de canales iónicos lo que se traduce en cambios en el potencial de reposo en una región dada o en toda la célula sensorial. Es el potencial receptor. Es un potencial local que a veces puede representar la única respuesta que resulta del proceso de transducción, por ejemplo en la célula sensorial gustativa.
Otras veces, este potencial es la respuesta al estímulo que se da en una región determinada de una célula, como es el caso del Corpúsculo de Pacini. Este es un terminal nervioso encapsulado que en respuesta a un estímulo mecánico genera un potencial local, graduado, que induce en el primer nódulo de Ranvier de ese axón, potenciales de acción propagados. A estos potenciales locales capaces de producir potenciales de acción se les llama también potenciales generadores.
En el modelo de transducción esquematizado en A se representa, en un segmento de la membrana de los cilios de la célula sensorial olfativa, un quimio-receptor de membrana al cual se unirá el estímulo (odógeno). Cuando éste se une al quimio-receptor, se activa la proteína G, lo cual induce la formación de segundos mensajeros, IP3, DG o cAMP, los cuales pueden modificar la permeabilidad iónica de la membrana, generando así un potencial receptor.
En el esquema B, se esquematiza un segmento de membrana del extremo terminal, encapsulado, de un corpúsculo de Pacini. El estímulo adecuado que actúa sobre este tipo de receptor, es un estímulo mecánico de presión, el cual deforma la cápsula que envuelve el terminal deformando también su membrana, lo que estimula elementos del citoesqueleto conectados a canales para el Na+, induciéndose así una despolarización local, el potencial receptor.
En el esquema C se muestra el proceso de transducción en la célula sensorial (cono o bastoncito) de un fotorreceptor de mamífero. El estímulo (energía de la luz) actúa sobre un pigmento (rodopsina) ubicado en un sistema de membranas internas (discos) de la célula receptora. Por este efecto se inicia una cadena de reacciones que lleva finalmente a la reducción de los niveles del segundo mensajero, el cGMP. En reposo, esta molécula es un activador de los canales de Na+, manteniéndolos abiertos, lo cual se traduce en que las células sensoriales estén depolarizadas en la obscuridad. Al disminuir el nivel del cGMP, se cierran los canales de Na+, produciéndose una hiperpolarización de la célula. Al dejar de influir ésta, inhibitoriamente sobre la célula bipolar, se inicia la activación de la vía.