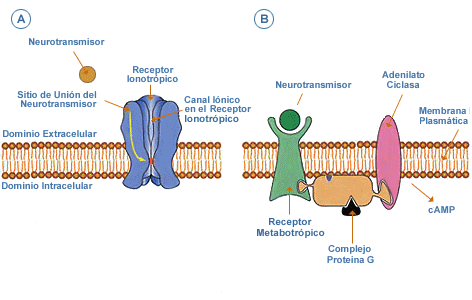


En la parte A del esquema se muestra un receptor-canal (ionotrópico), por ejemplo al neurotransmisor glutamato. Cuando este neurotransmisor se une a su sitio de unión en el receptor, se abre el canal iónico debido a una serie de cambios conformacionales en las subunidades que lo conforman. Este cambio permite la entrada del ión sodio, lo cual provoca una respuesta inmediata en ese punto, una hipopolarización, un PEPS.
El mismo neurotransmisor también se puede unir, simultáneamente, a otro subtipo de receptor, en este caso, metabotrópico. También se va a producir acá una serie de cambios conformacionales. Como resultado de esos cambios, el receptor puede influenciar a la proteína G, ubicada en su vecindad, lo cual provoca la activación de esa proteína.
La proteína G activada puede estimular a enzimas como la adenilato ciclasa, que induce la formación de un segundo mensajero, el adenosín monofosfato cíclico (cAMP). Esta molécula es capaz de activar enzimas dependientes de cAMP, como proteínas quinasas que catalizan la fosforilación de algunas subunidades del canal iónico. Ello provoca un cambio en la conductancia del canal, del receptor ionotrópipo, la cual puede disminuir.
La respuesta que se obtiene a través del receptor metabotrópico demora más en aparecer y es más duradera, desde décimas de segundo a horas.
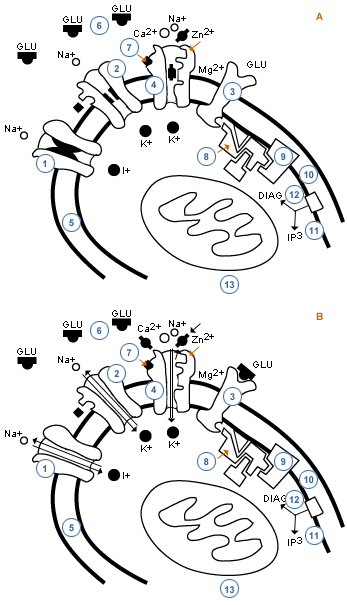
Tenemos entonces que entre los fenómenos más importantes que ocurren en las membranas, tanto pre como post-sinápticas, está el de las interacciones que se producen entre las respuestas que dan los distintos tipos de receptores al ser activados por sus respectivos neurotransmisores. También comentaremos las que pueden ocurrir entre los receptores ionotrópicos a un mismo neurotransmisor, como es el glutamato (o ácido glutámico).
En el esquema se muestran diversos subtipos de receptores a ese neurotransmisor. De ellos hay tres que corresponden a receptores ionotrópicos: a kainato, a AMPA y a NMDA. Los dos primeros subtipos de receptores son canales para el Na+ mientras que el tercero es un canal de Ca2+, pero que también permite el paso al K+ y al Na+. Este receptor-canal se encuentra normalmente bloqueado por el Mg2+, bloqueo que desaparece si la célula se hipopolariza ya que,en esas condiciones, se libera el ión magnesio del receptor.
Cuando en la sinapsis se libera el glutamato, este se unirá a los tres tipos de receptores con distinta velocidad y afinidad. Pero como resultado de la unión del glutamato a los receptores no-NMDA se produce, a través de ellos, entrada de Na+, se provoca en esa zona una hipopolarización. Cuando ésta alcanza una disminución de 20 a 30 mV, se sale el Mg2+ del receptor NMDA, el cual al estar también ocupado por glutamato se abre y permite la entrada de Ca2+.
Este receptor-canal sólo funciona en presencia de glicina (tiene sitio de unión a este aminoácido) y su activación se considera ligada a procesos tan importantes como la potenciación duradera (LTP), fenómeno ligado a su vez al proceso de la memoria.