|
|
 | |

|
|
| |
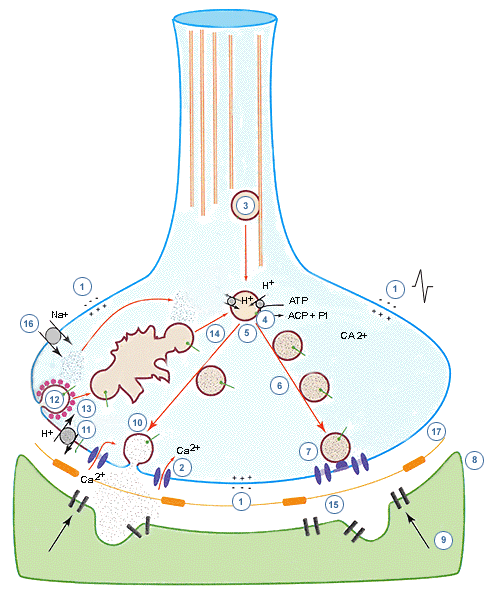
El mecanismo es muy complejo y en él juega un papel fundamental el Ca+2. Por la llegada del potencial de acción al terminal nervioso se abren los canales de calcio presentes en la membrana del terminal y el ión entra por difusión. Se produce así en la inmediata vecindad al interior de cada canal una momentánea alza de la concentración del ión.
Los canales se abren en el momento del pico del potencial de acción y el Ca+2 que entra genera un microdominio de elevada concentración del ión que está, sin embargo, a corta distancia del punto donde debe ejercere su efecto que es la vesícula ubicada en el sitio activo de liberación.
Normalmente en los sitios activos hay pocas vesículas ubicadas para entrar en el proceso de liberación. Por ello se le han asignado al calcio otras funciones además de la de iniciar la exocitosis. Una de ellas es la de acelerar el traslado de las vesículas a los sitios activos.
Pero además del calcio, tanto en el proceso de liberación como en el de traslado de vesículas juegan un papel importante componentes proteicos de las membranas vesicular y plasmática. Por una parte, las vesículas que no se encuentran en los sitios activos están agrupadas en el citoplasma presináptico, unidas entre sí o a la actina, gracias a una proteina, la sinapsina I, ubicada en la membrana de las vesículas. Cuando esta proteina es fosforilada se despegan las vesículas de los filamentos o de las otras vesículas. La fosforilación es catalizada por una quinasa II-dependiente de calmodulina que es, también, calcio-dependiente.: Esto significa que al entrar el ión calcio, se inicia el proceso de separación de vesículas del citoesqueleto para movilizarse a sitios activos. Como éstos se ubican alrrededor de los canales de entrada de calcio se producirá una rápida fusión de vesículas a la membrana del terminal.
ż Cómo se produce el proceso de ubicación de las vesículas en los sitios activos ? La ubicación se produce por un complicado proceso en que interactuan proteinas de la pared de la vesícula (sinaptobrevina y sinaptotagmina) y de la membrana del terminal (sintaxina y SNAP-25). Algunos de estos componentes se unen para formar un complejo llamado SNARE (sintaxina, SNAP-25 y sinaptobrevina) que interactua con dos proteinas solubles en el proceso de la ubicación: NSF (factor sensible a n-etilmaleimida) y a -SNAP, que es una proteina de unión a NSF, necesaria para que esta molécula (NSF) exprese su actividad ATP-ásica.
La ubicación de la vesículas en el sitio activo se explica, entonces, por la llamada hipótesis SNARE. Se asume que existe el complejo SNARE que al estar unido a la sinaptotagmina forma un complejo mayoor que une a la vesícula a un punto de la membrana. En esta situación, existe en la membrana vesicular una GTP-asa (hidroliza GTP) llamada Rab-3. Al hidrolizarse el GTP por la acción de esta enzima, el GDP resultante, se une a una proteina de la membrana plasmática (n-sec 1), fenómeno que permite la fusión de ambas membranas. Pero este fenómeno de fusión se iniciaría por la unión del calcio a la sinaptotagmina del complejo. Si no hay complejo SNARE la entrada del calcio no produciría la fusión.
Pero dadas las condiciones descritas, para que la fusión se produzca realmente, debe formarse previamente un poro de fusión, que es un poro del tamaño de una gap-junction, que se puede abrir o cerrar como un canal iónico. Cuando se abre este poro expandiéndose irreversiblemente se fusionan las membranas, pero al estar presente el poro quedan comunicados el inetrior de la vesícula con el espacio sináptico,
Luego sobreviene la exocitosis, es decir, la expulsión del contenido de la vesícula.